Mammalian Sex determination (HS)
Surprisingly, it is only in the last 50 years that we have begun to understand the nature of the biological events which determine our sex. It is not so long ago that women were blamed if they failed to produce a son for their husband and it was thought that the power of sex determination lay within the body of the woman. During this century the chromosomal basis of human sex determination has been demonstrated and in the last few years some of the genes responsible have been identified.
Sex chromosomes
The chromosomal basis of sex determination in humans was recognized when metaphase chromosomes from dividing male and female cells could be studied and counted. The normal karyotype contains 46 chromosomes including either two X chromosomes (46XX, females) or one X chromosome and one Y chromosome (46XY, males). This seems superficially similar to the situation in Drosophila melanogaster where females have two sets of autosomes and two X chromosomes (2AXX) whereas males are 2AXY. However we see a profound difference when we examine individuals with abnormal chromosome complements.
Karyotype |
Human |
Drosophila |
2A XX |
female |
female |
2A XY |
male |
male |
2A X0 |
female |
male |
2A XXX |
female |
female |
2A XXY |
male |
female |
Humans with 45X or 47XXX karyotypes are female and those with 47XXY karyotype are male. Therefore it can be deduced that the Y chromosome is sex determining in contrast to the Drosophila mechanism where it depends on the ratio of the number of X chromosomes to the number of haploid sets of autosomes.
Sex determination
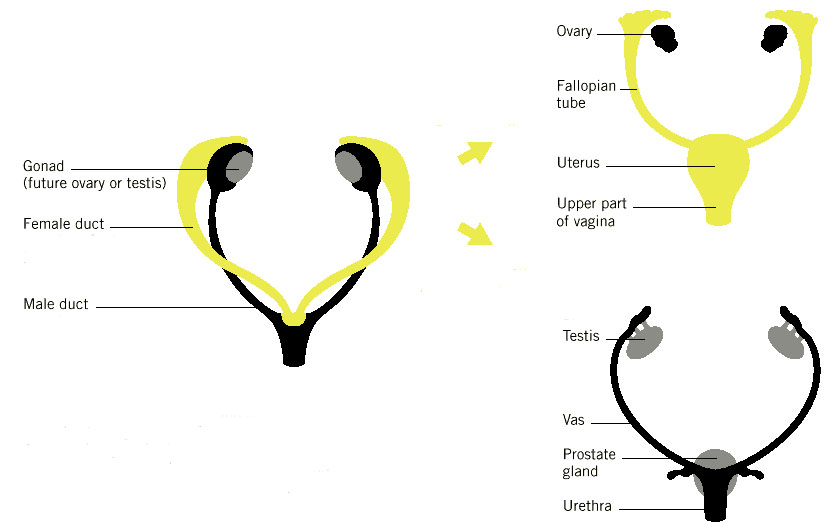
Experiments carried out by Jost in the 1960s involving the removal of the embryonic gonad have revealed that in mammals, no matter what the chromosomal sex of the somatic cells, the body will develop as a female unless a male gonad is present to secrete the hormones Mullerian inhibiting substance, MIS, (also known as Anti Mullerian Hormone, AMH) and testosterone. This can be partially mimicked in the genetic condition Androgen Insensitivity Syndrome, in which the gene coding for the androgen receptor is not expressed so that, although the testis in an XY individual secretes testosterone, the somatic tissues are unable to respond to it. Consequently the individual's body develops as a woman but with internal testes instead of ovaries.
Initially the gonad of both XX and XY embryos is bi-potential and both the Mullerian ducts and the Wolffian tubules are present. Once the male gonad begins to differentiate and secrete AMH and androgens the Mullerian duct regresses and the Wolffian duct develops into the vas deferens. In females the Mullerian duct persists and gives rise to the fallopian tubes.
Cells of the gonad
The embryonic gonad is comprised of four types of cell. In the mouse the structure is known as the germinal ridge. Initially the germ cells themselves are not present in the germinal ridge, they migrate to it from the yolk sac where they sought sanctuary during the upheaval of gastrulation. In mice homozygous for the mutation Steel this migration fails to occur, nevertheless the differentiation into testis in an XY individual still occurs. So germ cells are not necessary for correct differentiation. Transplantation experiments, in which male germ cells are transplanted to a female gonad, bear this out. The male germ cells in a female environment begin to develop as oocytes.
Candidate genes for TDF, the Testis Determining Factor
Studying XX males to work out what was the smallest region of the Y chromosome required for male development and screening this region for candidate genes led to the discovery of a gene coding for a protein with 13 zinc fingers named ZFY (Zinc Fingers, Y encoded).
Some factors which spoke against the role of ZFY in testes determiniation were:
Another gene was found named SRY (Sex Region on the Y chromosome) This gene is intronless. It has three domains, the N terminal domain, the so called Sry box, which resembles the DNA binding domain of the transcription factor HMG1, and a C-terminal domain. Several lines of evidence proved that SRY was a sex determining gene.
The human and mouse SRY genes are completely different except in the 79 amino acid SOX box which is 85% identical. Sry genes are found in all male mammals and there is no sequence conservation except in the SOX box. It is one of the most rapidly evolving genes known. Almost all mutations which cause XY females in humans are found in the SOX box. So most of the function is in that box. The human gene does not work in mice so the remaining structure may be of some importance.
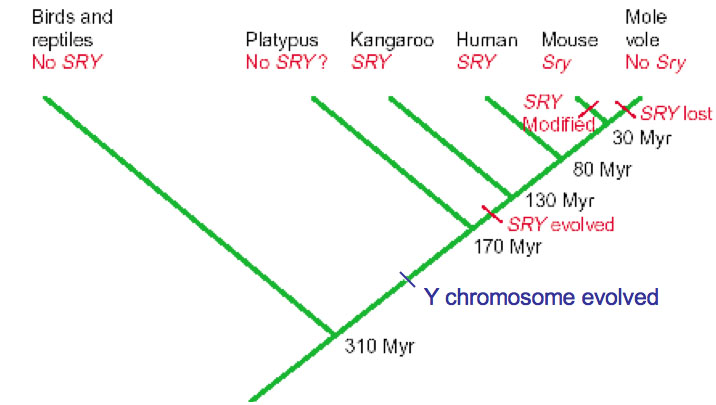
Sry is not present in monotremes and has been lost in some eutherians like the American mole vole. Since males of these species exist they presumably retain some other gene for testes determination.
SOX9
A disease which affects bone formation, campomelic dysplasia, is often accompanied by XY female sex reversal. Sox9 is expressed at 10.5 d.p.c. in the mouse in both sexes (shortly after the first appearance of Sry transcripts in males). It is then down regulated in females but its expression is increased in males. The gene is expressed in Sertoli cells throughout embryonic development and in adult males. Interestingly, it is also expressed in the developing Sertoli cells of male chickens (which are chromosomally ZZ and which don't have a recognisable SRY). Sox9 binds the promoter of the AMH gene in vitro. Other downstream targets (direct or indirect include SF1 and FGF9).
Ovary determining genes
Testes determining genes are either expressed exclusively in the male gonad or may (like Sox9) be transcribed in both sexes while the gonad is indifferent but be down regulated in female and upregulated in male gonads. Ovary determing genes should show the reverse pattern. One would also predict that loss of function mutations in such genes would have no effect in males but that gain of function mutations would lead to XY sex reversal.
Dax1, the mouse gene, is expressed in developing gonads. It seems to be repressed by Sry and in its turn perhaps to repress Sox9. Duplication of the chromosome band Xp21, which contains DAX1 gives rise to XY females. Deletion of this gene causes adrenal hypoplasia but no failure of ovary development in females. Genes which are required for ovary development and associated with at least partial sex reversal in XX mammals include the signaling factor Wnt4 and the forkhead protein FoxI2.
References
Gilbert 9th ed Chapter 17 pp 529-543
Ychromosome evolution
http://genome.wellcome.ac.uk/doc_WTD020741.html